NULL
Az ELTE és a HUN-REN Ökológiai Kutatóközpont Evolúciótudományi Intézetének (HUN-REN ÖK ETI) kutatói a többsejtűség kialakulását vizsgálták az általuk fejlesztett egyedalapú számítógépes modellben. Kétféle sejttípust modelleztek időben változó, térbeli környezetben. Az együttműködő sejtek aggregátumokat, összekapcsolódó sejtek halmazát hozhatják létre, míg a csalók nem termelik az aggregációhoz szükséges anyagot, de élvezik az aggregátum előnyeit. Az eredmények megerősítik, hogy forrásokban gazdag környezetben a kooperátorok lassabb növekedésük miatt hátrányban vannak. Ha azonban a források kifogynak, csak ezek tudnak termőtestet létrehozni és új élőhelyet meghódítani. A csalók csak az együttműködő sejteket kihasználva juthatnak el a következő, forrásokban gazdag élőhelyre. A PLOS Computational Biologyban megjelent eredmények igazolják, hogy a kooperátorok a térbeli aggregáció és a csoportszelekció révén változó környezetben is képesek túlélni minden további mechanizmus, például a ragadozók feltételezése nélkül.
Vajon hogyan és miért alakult ki a többsejtűség sokszor egymástól függetlenül az élővilág különböző ágain, és milyen tényezők járultak hozzá elterjedéséhez? Több elmélet is létezik, hogy miért előnyös többsejtűnek lenni. Az egysejtűek, például a baktériumok aggregációja szempontjából vannak nyilvánvalóan közvetlen előnnyel járó tényezők (például a ragadozók elkerülése), mások azonban, például a sejtek növekedésének térbeli mintázata és az élőhely időbeli változása, közvetett előnyökkel járnak. Ez utóbbiak ‒ csoportszelekcióval kapcsolva ‒ rokonfelismerés vagy ragadozó-nyomás nélkül is biztosíthatják a kooperatív (együttműködő sejtek) tulajdonság fennmaradását. A HUN-REN ÖK ETI és az ELTE kutatói egyedalapú számítógépes modelljükben ezt a hipotézist vizsgálták.
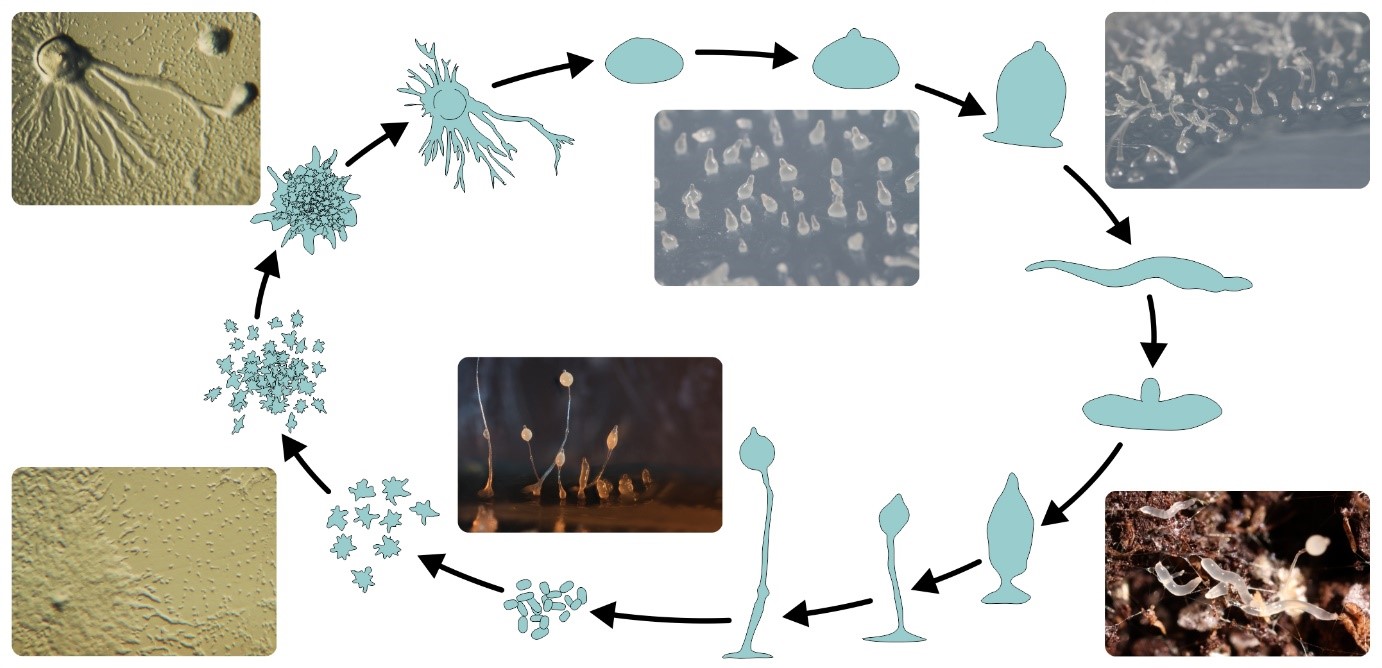
A többsejtűség kialakulása az evolúció egyik jelentős átmenete, amely több mint 25 alkalommal következett be az élővilág különböző ágain, egymástól függetlenül. Az összetett többsejtűek, mint amilyen az ember is, az osztódások során együttmaradó rokon sejtek révén érnek el nagyfokú bonyolultságot. Ezzel szemben a legtöbb egysejtű szervezet nem rendelkezik az ehhez szükséges szabályozási eszközökkel. Számukra többnyire csak egy egyszerűbb út jöhet számításba: időszakosan állnak össze többsejtű formává, például éhezés miatt. Ezek az aggregatív többsejtű fajok, mint például a Dictyostelium nyálkagomba, jellemzően egysejtű életet élnek. Ahogy azonban nevük is mutatja, a többsejtű, nyálkaszerű formák képesek mozogni, megfelelő termőhelyre eljutni és ott nyeles termőtestet növeszteni, amely által a spóráik messzebbre, táplálékgazdag új élőhelyekre is eljuthatnak.
Az ilyen többsejtűség esetében azonban nem-rokon egyedek, vagy akár olyanok is bekerülhetnek a túlélő sejtek közé, akik aktívan nem vesznek részt az együttműködésben, azaz csalnak. Mivel nem segítenek, minden energiájukat táplálkozásra és szaporodásra fordíthatják – az együttműködő sejtek kárára. Ezzel nemcsak az együttműködő sejtek túlélését veszélyeztetik, hanem végső soron a fajét is, hiszen ha túl sok a csaló, akkor nem lesz többsejtű forma, amely szaporíthatná a fajt. De hogyan tud akkor egy aggregatív többsejtű faj mégis túlélni, ha a csalók mindig gyorsabban szaporodnak az együttműködőknél? Különösen fontos kérdés ez az evolúciós átmenetek esetében, ahol kulcsfontosságú az együttműködés fenntartása a csalók ellenében.
Többféle elképzelés is van, amely magyarázatot adhat arra, miért látunk mégis sikeres aggregatív többsejtű fajokat. Az egyik elmélet szerint az aggregáció előnyös a ragadozókkal szembeni védekezésben: minél több egysejtű kapaszkodik össze, annál kevésbé tud egy mikrobiális ragadozó fogást találni rajtuk. Egy másik, szintén széles körben elfogadott feltételezés szerint a rendszeres éhezés miatt szükséges az új élőhelyek meghódítása, amihez nélkülözhetetlenek az együttműködő sejtek, így tehát a csalók is rájuk vannak utalva.
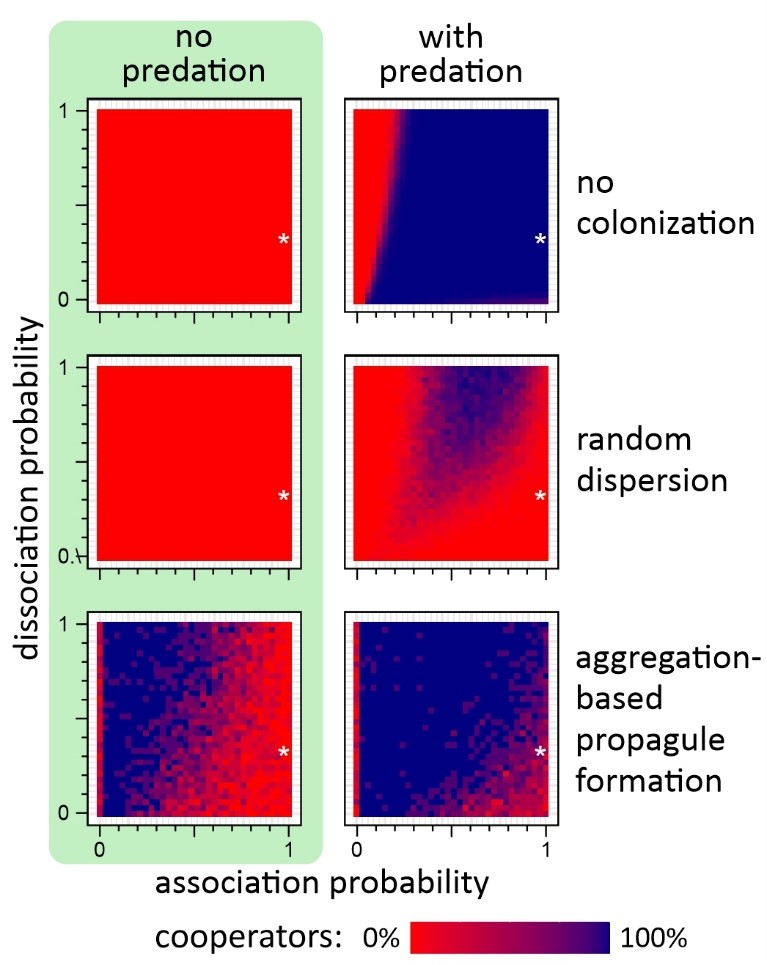
A HUN-REN ÖK ETI és az ELTE kutatói ezt a két feltételezést vizsgálták meg, azaz hogy milyen hatása van az aggregációnak és a kolonizációnak egyedi szelekció, illetve csoportszelekció esetén. Egyedalapú, térbeli számítógépes modellt fejlesztettek, amelyben egy nyálkagombaszerű egysejtű élőlény életciklusát modellezték. A modellben az együttműködő sejtek termelik az összeálláshoz szükséges „ragasztóanyagot”, a csalók nem. A számítógépes szimulációk egyértelműen igazolták, hogy időben folytonos környezetben, azaz ahol mindig van elegendő táplálék, a ragadozók elleni védekezés elengedhetetlen az együttműködő sejtek fennmaradásához. Azonban ha a környezetben időről időre forráshiány lép fel, a ragadozók okozta szelekció nemhogy nem elégséges, de nem is szükséges a kooperáció és a többsejtű forma fenntartásához – ekkor szükség van új élőhelyek kolonizálására.
A kutatók többféle kolonizációs mechanizmust is megvizsgáltak (diszperzió, fragmentáció, aggregatív spóraképzés stb.), és azt találták, hogy csak az aggregatív szaporító mechanizmusok képesek hosszú távon és robusztusan fenntartani a kooperációt ilyen változó környezetben. Vagyis változó környezetben a csoportszelekció fontosabb az egyedi szelekciós hatásoknál a kooperáció fenntartásában. Az eredmények azt sugallják, hogy ezeknek a mechanizmusoknak kulcsszerepe lehetett az aggregatív többsejtűek evolúciós kialakulásánál.