The nucleus may have been the necessary prerequisite/missing link for the acquisition of mitochondria by eukaryotes
NULL
This paper elaborates on the issues of microbial endosymbiosis stemming from conflict of interest and how multilevel selection can solve it once central control is gained by the host – given it has a nucleus. This is usually the case for extant endosymbiont of unicells (protists), where a huge asymmetry already exists (host being larger, with heterotrophic metabolism, assumed to be phagotrophic, having a nucleus, control mechanisms, etc.). However, among prokaryotes, this is not the case, as prokaryotic partners are far from being as different size- and structure-wise. Presumably there is something preventing prokaryotes to form endosymbiotic partnerships more often. The issue pertains especially to mitochondrial origins. Since we do not know if the ancestral host to mitochondria was nucleated or not, we cannot exclude the possibility that their success was not only due to the metabolic and architectural synergy the mitochondrial ancestor may have provided for the host but due to the fact that the host already could 1) shield its genome from uncontrolled hybridization with the symbiont’s genes plus 2) had the means to unilaterally exert control on the symbiont.
Our paper discusses why endosymbiosis (one microbe living inside another) is common in complex cells but rare in simple prokaryotes. In modern single-celled eukaryotes, the genetic interaction between host and symbiont is more asymmetric, potentially due to the nuclear envelope, acting as a barrier. In contrast, prokaryotes lack such features and are more equal, making stable partnerships harder. For mitochondria, it’s unclear if the original host had a nucleus, but if it did, that may have helped it control the symbiont and avoid harmful genetic interference – making the partnership more successful.
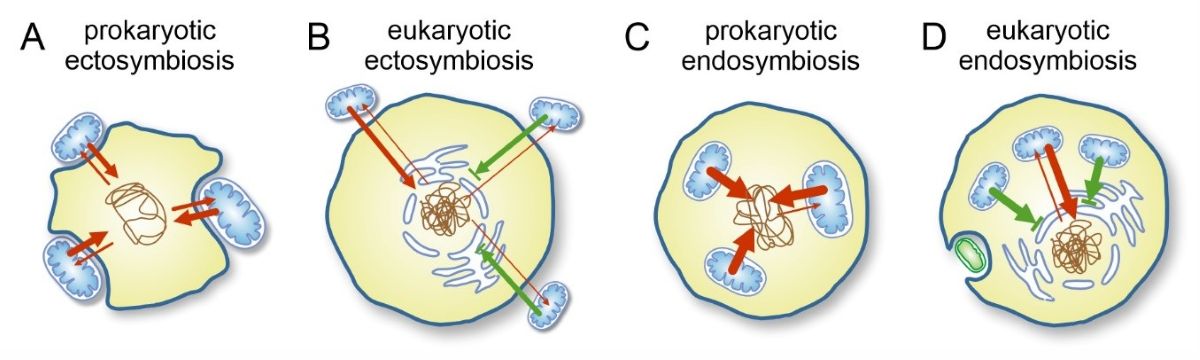
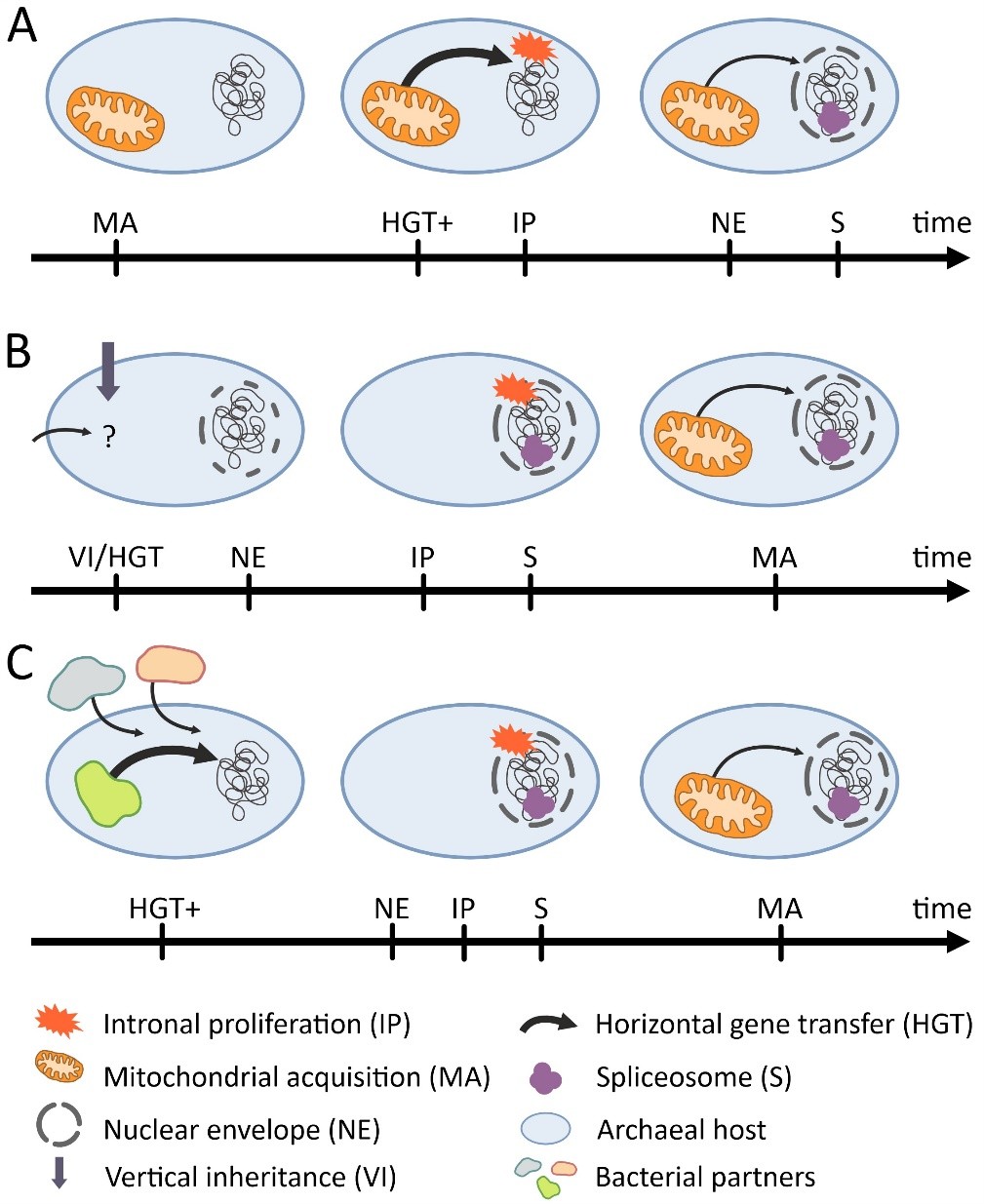
The amount of expected horizontal gene transfer (HGT) under various conditions of symbiosis. Red arrows indicate potential gene transfer, green arrows blocked transfer attempts; the strength of arrows indicates presumed magnitude of successful transfers in unit time.Overview of theories proposing various orders of events including the origin of nucleus, mitochondria and intronal evolution. A) mitochondria-early scenario of (Martin and Koonin 2006): An archaeon (without NE) acquires the mitochondrial endosymbiont as the first step. The endosymbiont incurs an increased level of HGT (EGT; thicker arrow) and as a result, Group II introns (among other genes) make their way into and proliferate in the host genome. Introns that lose their self-splicing ability become a burden for the host, and as a result, NE and spliceosomal machinery evolve only after intron proliferation. B) Early intron evolution and late mitochondrial acquisition based on (Vosseberg et al. 2020, 2022). Group II introns were either acquired by HGT by the host from pre-mitochondrial endosymbionts or were vertically inherited from archaeal introns already present in the lineage. A rudimentary version of NE is formed (or other mechanism to separate transcription and translation) with a wave of gene duplications potentially introducing further introns. As introns proliferated throughout the genome, the spliceosome evolved for the same reasons as described above. As the mitochondrial endosymbiont was established, a second wave of gene duplications and possibly more introns further shaped the proto-eukaryotic genome. C) Proposed hypothesis assuming serial endosymbiosis before mitochondria (Pittis and Gabaldón 2016, Gabaldón 2018), compatible with scenario B. An increased level of HGT from transient endosymbiotic partners perturb the host genome before the NE fully evolved. Early endosymbionts could not stabilize due to the lack of a NE that would enable controlled EGT and, ultimately, central control over the symbionts. The rest of this scenario follows B.